Learning Goals
By the end of this reading you should be able to:
- Describe the organizational features of the simplest multicellular animals
- Explain the various body forms and bodily functions of sponges
- Compare structural and organizational characteristics of Porifera and Cnidaria
- Describe the progressive development of tissues and their relevance to animal complexity
Introduction
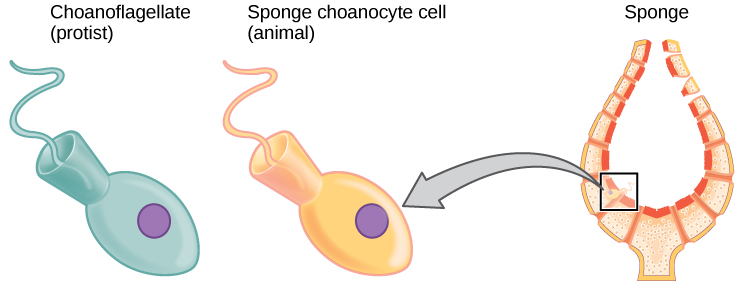
Biologists strive to understand the evolutionary history and relationships of members of the animal kingdom, and all of life, for that matter. Currently, most biologists divide the animal kingdom into 35 to 40 phyla. The current understanding of evolutionary relationships between animal, or Metazoa, phyla begins with the distinction between “true” animals with true differentiated tissues, called Eumetazoa, and animal phyla that do not have true differentiated tissues (such as the sponges), called Parazoa. Both Parazoa and Eumetazoa evolved from a common ancestral organism that resembles the modern-day protists called choanoflagellates (choan – Greek for “funnel”). These protist cells strongly resemble the sponge choanocyte cells today (Fig. 1).
Eumetazoa is subdivided into radially symmetrical animals and bilaterally symmetrical animals, and are thus classified into the clades Bilateria or Radiata, respectively. Cnidarians and ctenophores are animal phyla with true radial symmetry. All other Eumetazoa are members of the Bilateria clade. The bilaterally symmetrical animals are further divided into deuterostomes (including chordates and echinoderms) and two distinct clades of protostomes (including ecdysozoans and lophotrochozoans). Ecdysozoa includes nematodes and arthropods (Fig. 2); they are so named for a commonly found characteristic among the group: exoskeletal molting (termed ecdysis). Lophotrochozoa is named for two structural features, each common to certain phyla within the clade. Some lophotrochozoan phyla are characterized by a larval stage called trochophore larvae, and other phyla are characterized by the presence of a feeding structure called a lophophore.

Phylum Porifera
The invertebrates (Invertebrata) are animals that do not contain bony structures, such as a cranium and vertebrae. The simplest of all invertebrate animals are the Parazoans, which include only the phylum Porifera: the sponges. Parazoans do not display tissue-level organization, although they do have specialized cells that perform specific functions. While the larvae of sponges are able to swim the adults are non-motile (sessile) and spend their life attached to a solid surface. Since water is vital to sponges for excretion, feeding, and gas exchange, their body structure facilitates the movement of water through the sponge. Structures such as canals, chambers, and cavities enable water to move through the sponge to nearly all body cells.

Morphology of Sponges
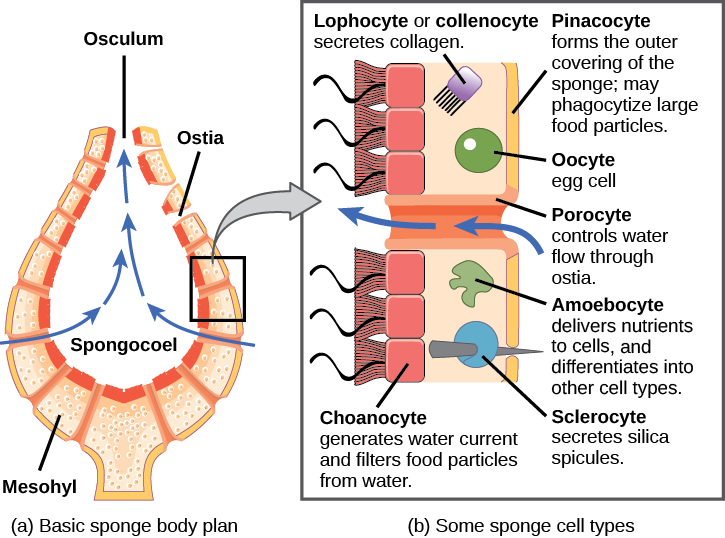
The morphology of the simplest sponges (Fig. 3) takes the shape of a cylinder with a large central cavity, the spongocoel, occupying the inside of the cylinder (Fig. 4). Water can enter into the spongocoel from numerous pores in the body wall. Water entering the spongocoel is expelled through a large common opening called the osculum. However, sponges exhibit a range of diversity in body forms, including variations in the size of the spongocoel, the number of osculi, and where the cells that filter food from the water are located. While sponges (excluding the hexactinellids) do not exhibit tissue-layer organization, they do have different cell types that perform distinct functions. Pinacocytes, which are epithelial-like cells, form the outermost layer of sponges and enclose a jelly-like substance called mesohyl. Mesohyl is an extracellular matrix consisting of a collagen-like gel with suspended cells that perform various functions. The gel-like consistency of mesohyl acts like an endoskeleton and maintains the tubular morphology of sponges. In addition to the osculum, sponges have multiple pores called ostia on their bodies that allow water to enter the sponge. In some sponges, ostia are formed by porocytes, single tube-shaped cells that act as valves to regulate the flow of water into the spongocoel. In other sponges, ostia are formed by folds in the body wall of the sponge. Choanocytes (“collar cells”) are present at various locations, depending on the type of sponge, but they always line the inner portions of some space through which water flows (the spongocoel in simple sponges, canals within the body wall in more complex sponges, and chambers scattered throughout the body in the most complex sponges). Whereas pinacocytes line the outside of the sponge, choanocytes tend to line certain inner portions of the sponge body that surround the mesohyl. The structure of a choanocyte is critical to its function, which is to generate a water current through the sponge and to trap and ingest food particles by phagocytosis. Note the similarity in appearance between the sponge choanocyte and choanoflagellates (Protista). This similarity suggests that sponges and choanoflagellates are closely related and likely share a recent common ancestry. The cell body is embedded in mesohyl and contains all organelles required for normal cell function, but protruding into the “open space” inside of the sponge is a mesh-like collar composed of microvilli with a single flagellum in the center of the column. The cumulative effect of the flagella from all choanocytes aids the movement of water through the sponge: drawing water into the sponge through the numerous ostia, into the spaces lined by choanocytes, and eventually out through the osculum (or osculi). In the meantime, food particles, including waterborne bacteria and algae, are trapped by the sieve-like collar of the choanocytes, slide down into the body of the cell, are ingested by phagocytosis, and become encased in a food vacuole. Lastly, choanocytes will differentiate into sperm for sexual reproduction, where they will become dislodged from the mesohyl and leave the sponge with expelled water through the osculum.
Watch this video to see the movement of water through the sponge body. https://youtu.be/pTZ211cIjX8
The second crucial cells in sponges are called amoebocytes, named for the fact that they move throughout the mesohyl in an amoeba-like fashion. Amoebocytes have a variety of functions: delivering nutrients from choanocytes to other cells within the sponge, giving rise to eggs for sexual reproduction (which remain in the mesohyl), delivering phagocytized sperm from choanocytes to eggs, and differentiating into more-specific cell types. Some of these more specific cell types include collencytes and lophocytes, which produce the collagen-like protein to maintain the mesohyl, sclerocytes, which produce spicules in some sponges, and spongocytes, which produce the protein spongin in the majority of sponges. These cells produce collagen to maintain the consistency of the mesohyl.
Review Question:
Sponges
In some sponges, sclerocytes secrete small spicules into the mesohyl, which are composed of either calcium carbonate or silica, depending on the type of sponge. These spicules serve to provide additional stiffness to the body of the sponge. Additionally, spicules, when present externally, may ward off predators. Another type of protein, spongin, may also be present in the mesohyl of some sponges.
Physiological Processes in Sponges
Sponges, despite being simple organisms, regulate their different physiological processes through a variety of mechanisms. These processes regulate their metabolism, reproduction, and locomotion.
Digestion: Sponges lack complex digestive, respiratory, circulatory, reproductive, and nervous systems. Their food is trapped when water passes through the ostia and out through the osculum. Bacteria smaller than 0.5 microns in size are trapped by choanocytes, which are the principal cells engaged in nutrition, and are ingested by phagocytosis. Particles that are larger than the ostia may be phagocytized by pinacocytes. In some sponges, amoebocytes transport food from cells that have ingested food particles to those that do not. For this type of digestion, in which food particles are digested within individual cells, the sponge draws water through diffusion. The limit of this type of digestion is that food particles must be smaller than individual cells. All other major body functions in the sponge (gas exchange, circulation, excretion) are performed by diffusion between the cells that line the openings within the sponge and the water that is passing through those openings. All cell types within the sponge obtain oxygen from water through diffusion. Likewise, carbon dioxide is released into seawater by diffusion. In addition, nitrogenous waste produced as a byproduct of protein metabolism is excreted via diffusion by individual cells into the water as it passes through the sponge.
Reproduction: Sponges reproduce by sexual as well as asexual methods. The typical means of asexual reproduction is either fragmentation (where a piece of the sponge breaks off, settles on a new substrate, and develops into a new individual) or budding (a genetically identical outgrowth grows from the parent and eventually detaches or remains attached to form a colony). An atypical type of asexual reproduction is found only in freshwater sponges and occurs through the formation of gemmules. Gemmules are environmentally resistant structures produced by adult sponges wherein the typical sponge morphology is inverted. In gemmules, an inner layer of amoebocytes is surrounded by a layer of collagen (spongin) that may be reinforced by spicules. The collagen that is normally found in the mesohyl becomes the outer protective layer. In freshwater sponges, gemmules may survive hostile environmental conditions like changes in temperature and serve to recolonize the habitat once environmental conditions stabilize. Gemmules are capable of attaching to a substratum and generating a new sponge. Since gemmules can withstand harsh environments, are resistant to desiccation, and remain dormant for long periods, they are an excellent means of colonization for a sessile organism.
Sexual reproduction in sponges occurs when gametes are generated. Sponges are monoecious (hermaphroditic), which means that one individual can produce both gametes (eggs and sperm) simultaneously. In some sponges, the production of gametes may occur throughout the year, whereas other sponges may show sexual cycles depending upon water temperature. Sponges may also become sequentially hermaphroditic, producing oocytes first and spermatozoa later. Oocytes arise by the differentiation of amoebocytes and are retained within the spongocoel, whereas spermatozoa result from the differentiation of choanocytes and are ejected via the osculum. Ejection of spermatozoa may be a timed and coordinated event, as seen in certain species. Spermatozoa carried along by water currents can fertilize the oocytes borne in the mesohyl of other sponges. Early larval development occurs within the sponge, and free-swimming larvae are then released via the osculum.
Locomotion: Sponges are generally sessile as adults and spend their lives attached to a fixed substratum. They do not show movement over large distances like other free-swimming marine invertebrates. However, sponge cells are capable of creeping along substrata via organizational plasticity. Under experimental conditions, researchers have shown that sponge cells spread on physical support demonstrate a leading edge for directed movement. It has been speculated that this localized creeping movement may help sponges adjust to microenvironments near the point of attachment. It must be noted, however, that this pattern of movement has been documented in laboratories, but it remains to be observed in natural sponge habitats.
Phylum Cnidaria
The Phylum Cnidaria includes animals that show radial or biradial symmetry and are diploblastic, that is, they develop from two embryonic layers. Nearly all (about 99 percent) cnidarians are marine species.

Cnidarians contain specialized cells known as cnidocytes (“stinging cells”) with organelles called nematocysts that contain coiled threads that may bear barb (stingers) (Fig. 5). These cells are present around the mouth and tentacles. The outer wall of the cell has hairlike projections, which are sensitive to touch. When touched, the cells are known to fire coiled threads that can either penetrate the flesh of the prey or predators of cnidarians or ensnare it. These coiled threads release toxins into the target and can often immobilize prey or scare away predators.

Cnidarians display two distinct morphological body plans: polyp or “stalk” and medusa or “bell” (Fig. 6). An example of the polyp form is Hydra spp.; perhaps the most well-known medusoid animals are the jellies (jellyfish). Polyp forms are sessile as adults, with a single opening to the digestive system (the mouth) facing up with tentacles surrounding it. Medusa forms are motile, with the mouth and tentacles hanging down from an umbrella-shaped bell. Some cnidarians are polymorphic, that is, they have two body plans during their life cycle.
All cnidarians show the presence of two membrane layers in the body that are derived from the endoderm and ectoderm of the embryo. The outer layer (from ectoderm) is called the epidermis and lines the outside of the animal, whereas the inner layer (from endoderm) is called the gastrodermis and lines the digestive cavity. Between these two membrane layers is a non-living, jelly-like mesoglea connective layer. In terms of cellular complexity, cnidarians show the presence of differentiated cell types in each tissue layer, such as nerve cells, contractile epithelial cells, enzyme-secreting cells, and nutrient-absorbing cells, as well as the presence of intercellular connections. However, the development of organs or organ systems is not advanced in this phylum. The nervous system is primitive, with nerve cells scattered across the body.
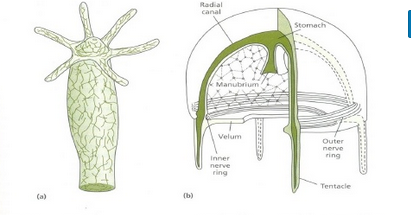
The nerve cells show mixed characteristics of motor as well as sensory neurons (Fig. 7). The predominant signaling molecules in these primitive nervous systems are chemical peptides, which perform both excitatory and inhibitory functions. Despite the simplicity of the nervous system, it coordinates the movement of tentacles, the drawing of captured prey to the mouth, the digestion of food, and the expulsion of waste.
The cnidarians perform extracellular digestion in which the food is taken into the gastrovascular cavity, enzymes are secreted into the cavity, and the cells lining the cavity absorb nutrients. The gastrovascular cavity has only one opening that serves as both a mouth and an anus, which is termed an incomplete digestive system.
Cnidarian cells exchange oxygen and carbon dioxide by diffusion between cells in the epidermis with water in the environment, and between cells in the gastrodermis with water in the gastrovascular cavity. The lack of a circulatory system to move dissolved gases limits the thickness of the body wall and necessitates a non-living mesoglea between the layers. There is no excretory system or organs, and nitrogenous wastes simply diffuse from the cells into the water outside the animal or in the gastrovascular cavity. There is also no circulatory system, so nutrients must move from the cells that absorb them in the lining of the gastrovascular cavity through the mesoglea to other cells.
Summary
Animals included in phylum Porifera are parazoans because they do not show the formation of true embryonically derived tissues, although they have a number of specific cell types and “functional” tissues such as pinacoderm. These organisms show very simple organization, with a rudimentary endoskeleton of spicules and spongin fibers. Glass sponge cells are connected together in a multinucleated syncytium. Although sponges are very simple in organization, they perform most of the physiological functions typical of more complex animals.
Cnidarians represent a more complex level of organization than Porifera. They possess outer and inner tissue layers that sandwich a noncellular mesoglea between them. Cnidarians possess a well-formed digestive system and carry out extracellular digestion in a digestive cavity that extends through much of the animal. The mouth is surrounded by tentacles that contain large numbers of cnidocytes—specialized cells bearing nematocysts used for stinging and capturing prey as well as discouraging predators. Cnidarians have separate sexes and many have a lifecycle that involves two distinct morphological forms—medusoid and polypoid—at various stages in their life cycles. In species with both forms, the medusa is the sexual, gamete-producing stage and the polyp is the asexual stage. Cnidarian species include individual or colonial polypoid forms, floating colonies, or large individual medusa forms (sea jellies).
End of Section Review Questions:
Review: Sponges
Review: Sponge amoebocytes
Review: Porifera vs Cnidaria
B
1,A; 2,C; 3,B
A, C, D
A, B, C
A, C, D
