4
Muistin on ajateltu jo pitkään olevan jaettavissa kolmeen alalajiin. Kognitiiviset (aivo)tieteilijät toki kehittävät tätä perinteistä mallia edelleen. Mallissa muistin kolme alalajia ovat (1) sensorinen muisti, (2) lyhytkestoinen (työ)muisti ja (3) säilömuisti, jota kutsutaan toisinaan myös pitkäkestoiseksi muistiksi.
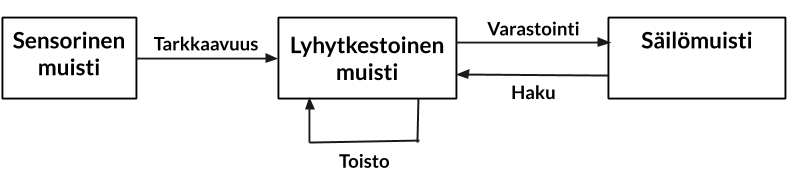
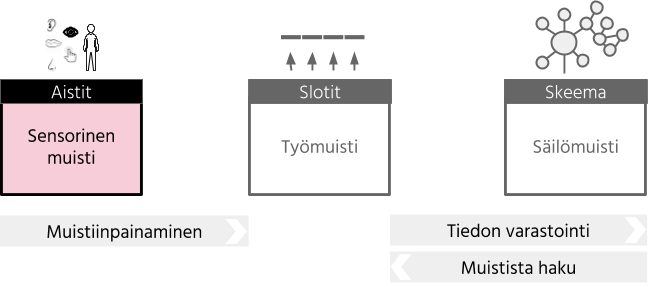
Sensorinen muisti on havaitsemiseen liittyvä automaattinen prosessi. “Sensorisella muistilla viitataan automaattiseen ja tilapäiseen aistiärsykkeistä saatavan tiedon säilömiseen, mikä mahdollistaa saadun tiedon yhdistämisen edellisiin ärsykkeihin ja/tai muistista haettuun tietoon” (Alain, Woods & Knight, 1998). Aistien avulla kerätään jatkuvasti tietoa ympäristöstä ja tämä tieto on aina saatavilla. Kun tarkkaavuus (huomio) suunnataan tai se tahattomasti suuntautuu tiettyyn aistiin, tulemme tietoiseksi aistin vastaanottamasta tiedosta. Tämä tietoisuus on merkki siitä, että aistitieto on siirtynyt työmuistiimme, joka on osa lyhytkestoista muistia. Tarkkaavuus voidaan siirtää milloin tahansa tietyn aistin kautta saapuvaan tietoon, jotta tätä aistitietoa voidaan tarkemmin tarkastella. Lyhytkestoisen muistin voidaan ajatella kuvastavan niitä ihmismielen kykyjä, jotka hetkellisesti ylläpitävät rajallista tietomäärää helposti saatavilla olevassa muodossa (Cowan, 2008).
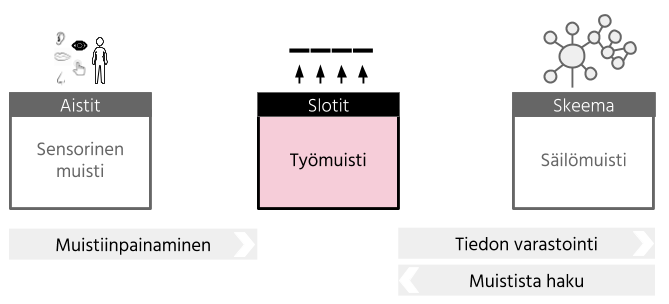
Työmuistin taas on kuvattu olevan lyhytkestoista tiedon ylläpitoa ilman aistitiedon syötettä (Eriksson, Vogel, Lansner, Bergström & Nyberg, 2015). Työmuisti sisältää lyhytkestoisen muistin lisäksi prosessointimekanismeja, joilla lyhytkestoista muistia voidaan hyödyntää (Cowan, 2008). Työmuistin ajatellaankin olevan lyhytkestoisen muistin osa, jossa aktiivisesti ylläpidetään ja muokataan lyhytkestoisessa muistissa olevaa tietoa.
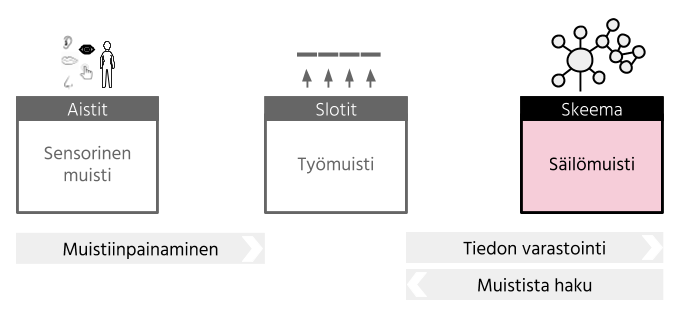
Säilömuisti tai pitkäkestoinen muisti (engl. long-term memory) taas voidaan ajatella valtavana tiedon varastona ja aiempien tapahtumien tallenteena (Cowan, 2008). Säilömuistin ja lyhytkestoisen muistin on esitetty eroavan kahdella perustavanlaatuisella tavalla toisistaan: vain lyhytkestoisessa muistissa muistiaines hiipuu ajan myötä ja sen kapasiteetti on rajallinen. (Cowan, 2008). Säilömuistin muistikuvien heikkeneminen ei taas tapahdu yhtä nopeasti kuin lyhytkestoisessa muistissa, eikä säilömuistiin varastoidulle tiedon määrälle voida määrittää selkeää rajaa.
Lyhytkestoisen muistin ja säilömuistin jako toisistaan erillisiksi järjestelmiksi on todennettu eri tutkimuksissa. Muistijäljen muodostuminen joko lyhytkestoisessa muistissa tai säilömuistissa on kyetty tutkimuksissa estämään ilman vaikutusta toiseen muistiin muodostuvaan muistijälkeen (Cammarota, M., Bevilaqua, Medina & Izquierdo, 2007). Esimerkiksi estämällä tiettyjen aivojen välittäjäaineiden reseptoreiden toimintaa lyhytkestoisen muistijäljen muodostuminen häiriintyy, mutta säilömuistiin muistijälki kuitenkin tallentuu. Vastaavasti häiritsemällä proteiinisynteesiä lyhytkestoinen muistijälki muodostuu, mutta muistijäljen muodostuminen ja päivittyminen säilömuistissa estyy. Myös erilaiset muistin menetykset eli amnesiat tukevat käsitystä erillisistä muistin alalajeista (Squire ym., 2015). Kyvyttömyys luoda uusia muistikuvia amnesian aiheuttaneen tapahtuman jälkeen, eli anterogradinen amnesia, johtaa osittaiseen tai täyteen kyvyttömyyteen muistaa lähimenneisyyttä. Tästä huolimatta tapahtumaa edeltäneet säilömuistin muistot ovat vahingoittumattomia. Vastakohtana tälle on retrogradinen amnesia, jossa amnesian aiheuttamaa tapahtumaa edeltävät muistikuvat katoavat, mutta uusia muistikuvia voidaan tapahtuman jälkeen muodostaa.
Aivojen erillisten muistijärjestelmien ajatellaan palvelevan selviytymisen kannalta eri tarkoituksia (Fuster & Bressler, 2015). Lyhytkestoinen muisti mahdollistaa elintärkeän mukautumisen uusiin tilanteisiin ja uuteen tietoon jatkuvasti muuttuvassa ja epävarmassa ympäristössä. Säilömuisti taas mahdollistaa tehokkaan varautumisen tuleviin kokemuksiin. Muistin biologisen hyödyllisyyden ajatellaankin perustuvan kykyyn muokata tulevaa käyttäytymistä aiempien kokemusten perusteella (Kukushkin & Carew, 2017). Sen avulla on mahdollista tunnistaa elinympäristössä toistuvia tilanteita ja reagoida niihin entistä tehokkaammin (Schacter, Addis & Buckner, 2007).
Taulukko 1. Lyhytkestoisen muistin ja sälömuistin tunnuspiirteet1
| Tunnuspiirteet | Lyhytkestoinen (työ)muisti | Säilömuisti |
| Kesto | 10-15 s ilman toistoa | Minuuteista elinikään |
| Kapasiteetti | 4 yksikköä tai tietokokonaisuutta | Ei tunnettua rajaa |
| Muistikuvien muodostumisen nopeus | Nopeaa, sekunnin tuhannesosia | Hidasta, minuuteista päiviin |
| Aivoalue | Aivoturso eli hippokampus | Levittäytyneenä aivokuoren uloimmalle osalle |
| Mekanismi | Aivojen välittäjäaineet | Uusien proteiinien synteesi, geeniekspressio |
1 Kukushkin & Carew, 2017; Eriksson ym., 2015; Kandel, Dudai, & Mayford, 2014
Lähteet: Osa 1
Muistin alalajit
- Fuster, J. M., & Bressler, S. L. (2015). Past makes future: role of pFC in prediction. Journal of cognitive neuroscience, 27(4), 639-654.
- Fuster, J. M. (2009). Cortex and memory: emergence of a new paradigm. Journal of cognitive neuroscience, 21(11), 2047-2072.
- Squire, L. R., Genzel, L., Wixted, J. T., & Morris, R. G. (2015). Memory consolidation. Cold Spring Harbor perspectives in biology, 7(8), a021766.
- Hunkin, N., Parkin, A., Bradley, V., Burrows, E., Aldrich, F., Jansari, A., & Burdon-Cooper, C. (1995) Focal retrograde amnesia following closed head injury: A case study and theoretical account, Neuropsychologia, 33(4) 509-523. doi:10.1016/0028-3932(94)00136-D
- Schacter, D. L., Addis, D. R., & Buckner, R. L. (2007). Remembering the past to imagine the future: the prospective brain. Nature reviews neuroscience, 8(9), 657-661.
- Kukushkin, N. V., & Carew, T. J. (2017). Memory Takes Time. Neuron, 95(2), 259-279.
- Cowan, N. (2008). What are the differences between long-term, short-term, and working memory?. Progress in brain research, 169, 323-338.
- Alain, C., Woods, D. L., & Knight, R. T. (1998). A distributed cortical network for auditory sensory memory in humans. Brain research, 812(1-2), 23-37.
- Eriksson, J., Vogel, E. K., Lansner, A., Bergström, F., & Nyberg, L. (2015). Neurocognitive architecture of working memory. Neuron, 88(1), 33-46.
- Kandel, E. R., Dudai, Y., & Mayford, M. R. (2014). The molecular and systems biology of memory. Cell, 157(1), 163-186.
- Cammarota, M., Bevilaqua, L. R., Medina, J. H., & Izquierdo, I. (2007). 10 Studies of Short-Term Avoidance Memory. Neural plasticity and memory: from genes to brain imaging, 193.

{kind=link}